Ecological and Evolutionary Dynamics
in Lakes, Bottles and Models
Research Approaches, Projects and Sites
I have been interested in ways that organisms respond ecologically and evolutionarily to variable environments. Variation in time and space may result from changes in the external environment, or from the interactions among individuals and species within the community. Evolutionary, population and community responses to environmental change (whether natural or caused by humans), are in large part a function of the diversity of genotypes and species present and their differing levels of success. Together with my close Cornell colleague, theoretician Steve Ellner, and many great postdocs and students, we have found that rapid evolution on the same time scale as ecological dynamics can dramatically alter ecological patterns and processes while they are occurring – a process known as “eco-evolutionary dynamics.” We’ve shown effects on predator-prey cycles, seasonal plankton dynamics, and the long-term persistence of genetic variation. My group has also shown that zooplankton make dormant eggs that can survive in lake sediments for decades or centuries and then hatch, affecting population evolutionary potential. These dormant egg banks are also a resource for studying past evolutionary events since viable eggs can be hatched from different times in the past. With colleagues at the former Max-Planck-Institute for Limnology (now MPI-Evolutionary Biology) we’ve used the egg bank in Lake Constance to show how Daphnia evolved resistance to toxic cyanobacteria as these primary producers increased in abundance during eutrophication. I am currently studying how climate change (both warming and increased storm frequency) is affecting seasonal thermal stratification and mixing patterns in lakes and how that influences plankton dynamics, especially the formation of harmful cyanobacterial blooms, which can feedback as natural selection on Daphnia dietary resistance.
Since moving to Hawaii Island, I have continued studies of the ecology of the endemic shrimp, Halocardina rubra, in anchialine pools along the Kona-Kohala Coast. I have also initiated a study of the factors influencing the success of a highly invasive freshwater copepod, Arctodiaptomus dorsalis, native to countries around the Caribbean Sea. It has invaded lakes in the Philippines and I found it dominant on Hawaii Island in the lake in Kapoho Crater before it filled with lava during the 2018 eruption of Kilauea volcano. It is currently dominant in farm ponds containing fish to the exclusion of virtually all other zooplankton. I have found that this tropical diaptomid copepod makes diapausing eggs which may explain its ability to disperse.
Since moving to Hawaii Island, I have continued studies of the ecology of the endemic shrimp, Halocardina rubra, in anchialine pools along the Kona-Kohala Coast. I have also initiated a study of the factors influencing the success of a highly invasive freshwater copepod, Arctodiaptomus dorsalis, native to countries around the Caribbean Sea. It has invaded lakes in the Philippines and I found it dominant on Hawaii Island in the lake in Kapoho Crater before it filled with lava during the 2018 eruption of Kilauea volcano. It is currently dominant in farm ponds containing fish to the exclusion of virtually all other zooplankton. I have found that this tropical diaptomid copepod makes diapausing eggs which may explain its ability to disperse.

NOTE PLEASE THAT I RETIRED IN DECEMBER 2019 AND AM NOT ACCEPTING ANY NEW LAB MEMBERS,
INCLUDING POSTDOCS, GRAD STUDENTS AND UNDERGRADS.
ABOUT OUR STUDENTS
Graduate students in my lab have worked on ecological and evolutionary processes in aquatic organisms (usually in fresh water), but the topics range broadly. We all learn from each other when the breadth of research topics explored by students is wide. In some cases students are co-advised by colleagues. In the past decade or so, my PhD students have investigated (1) the environmental and trait determinants of the relationship between local abundance and species range, especially in freshwater copepods (Rachel Wilkins), (2)
the positive and negative roles of colonial cyanobacteria on pelagic community and ecosystem processes in oligotrophic lakes (Cayelan Carey), (3) the effect of plankton and predator dispersal on plankton metacommunity (trophic cascade) dynamics in a system of island rock pools (Juniper Simonis), (4) the relative importance of autochthonous and allochthonous primary and heterotrophic production on stream communities (Sarah Collins; co-advised by Alex Flecker), (5) the indirect effects of fish adaptive morphologies in response to predation intensity on nutrient cycling (Chris Dalton; co-advised by Alex Flecker), (6) the role of native fishes in sediment processing and transport in desert streams (Mike Booth; co-advised by Alex Flecker), (7) the relative importance of external forcing versus interspecific interactions in determining plankton population dynamics in embayments along L. Ontario (Becky Doyle Morin), (8) the spatial and temporal population structure of bioluminescent marine ostracods (Gretchen Gerrish; co-advised by Jim Morin), (9) the microevolutionary response of Daphnia to rapid environmental change in a polluted lake (Jennifer Fox), and (10) the factors affecting food chain length in lakes of differing size and productivity (David Post; co-advised by Mike Pace).
Undergraduate students have worked in my lab either on my research projects (or those of my grad students and postdocs), or carry out their own independent studies. Independent investigations have involved (1) genes involved in DNA repair after UV damage in high mountain lake Daphnia (Paige Kulling) (2) predatory-prey dynamics in lab microcosms and theory by tuning the strength of rotifer predation on algae using salinity (Alex Looi, Kathryn Blackley, Dave Rosenberg), (3) strength of evolutionary rescue as a function of extent of inbreeding depression in rotifers (Kathryn Blackley), (4) rates of plankton dispersal among rock pools during rain-induced overflow events (Kara Pellowe), (5) tracing clonal lineages of algae using microsatellite-DNA to follow evolutionary response of algae to consumption by rotifers (Justin Meyer), (6) the patterns of Daphnia abundance in Austrian Alpine lakes with very long histories of fish introduction (Linda Perry Demma), (7) the identification of past invasions by exotic species in using molecular analysis of dormant eggs from lake sediments (Torrie Hanley), (8) molecular genetic analysis of genetic variation in copepod species with and without prolonged diapause, and genetic changes in algal prey during predator-prey cycles (Matt Holland). When appropriate, undergraduate students participate as co-authors on publications (e.g., Hewson, NG, Li, Aguirre, Looi et al. 2013; Meyer et al. 2006; Bohonak, Holland et al. 2006; Hairston...Perry et al. 2005; Duffy, Perry et al. 2000; Hairston, Perry et al. 1999 - all names except mine, Hewson’s, and Bohonak's were undergrads in my lab when the published work was carried out).
PAST RESEARCH
Eco-evolutionary dynamics and internally-driven temporal variation – Along with Stephen Ellner, Laura Jones (Cornell University), and a series of great postdocs (Gregor Fussmann, now on the faculty at McGill University; Takehito Yoshida, now on the faculty at Tokyo University; Lutz Becks, on the faculty at the University of Konstanz; Teppo Hiltunen, now on the faculty at the University of Turku; Brooks Miner, now on the faculty at Ithaca College, and Masato Yamamichi, now on the faculty at the University of Queensland), as well as a cadre of dedicated undergraduate students, we used highly controlled culture conditions to create predator-prey oscillations between algae and rotifers. We were able to predict the dynamics in our culture system with mechanistic mathematical models (Fussmann et al. 2000) and then to use these models to predict unexpected behaviors. One very interesting result was the observation that when the prey (algae) can evolve in response to temporally varying selection imposed by predation (rotifer grazing) and competition for limiting nutrients (nitrogen), the pattern of the predator-prey oscillations is radically altered (Yoshida et al. 2003). Under some conditions, "cryptic oscillations" of prey genotypes occur while total prey density remains essentially constant (Yoshida et al. 2007). Thus rapid contemporary evolution can dramatically alter what has traditionally been viewed as a purely ecological process (Becks et al. 2010). We have recently used this system to study the functional genomics of oscillating evolution of prey defense (cell clumping by Chlamydomonas) in response to predator-prey cycling (Becks et al. 2012).
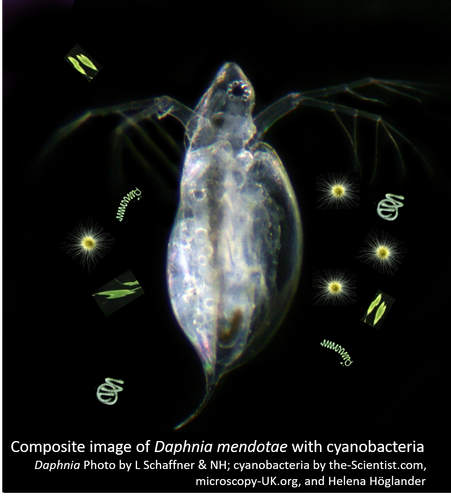
In 2019, we published a paper (Schaffner & Govaert et al. 2019) showing that Daphnia mendotae evolves resistance to summer cyanobacterial blooms within a single season and that this evolution feeds back to affect the population dynamics of the Daphnia.
INCLUDING POSTDOCS, GRAD STUDENTS AND UNDERGRADS.
ABOUT OUR STUDENTS
Graduate students in my lab have worked on ecological and evolutionary processes in aquatic organisms (usually in fresh water), but the topics range broadly. We all learn from each other when the breadth of research topics explored by students is wide. In some cases students are co-advised by colleagues. In the past decade or so, my PhD students have investigated (1) the environmental and trait determinants of the relationship between local abundance and species range, especially in freshwater copepods (Rachel Wilkins), (2)
the positive and negative roles of colonial cyanobacteria on pelagic community and ecosystem processes in oligotrophic lakes (Cayelan Carey), (3) the effect of plankton and predator dispersal on plankton metacommunity (trophic cascade) dynamics in a system of island rock pools (Juniper Simonis), (4) the relative importance of autochthonous and allochthonous primary and heterotrophic production on stream communities (Sarah Collins; co-advised by Alex Flecker), (5) the indirect effects of fish adaptive morphologies in response to predation intensity on nutrient cycling (Chris Dalton; co-advised by Alex Flecker), (6) the role of native fishes in sediment processing and transport in desert streams (Mike Booth; co-advised by Alex Flecker), (7) the relative importance of external forcing versus interspecific interactions in determining plankton population dynamics in embayments along L. Ontario (Becky Doyle Morin), (8) the spatial and temporal population structure of bioluminescent marine ostracods (Gretchen Gerrish; co-advised by Jim Morin), (9) the microevolutionary response of Daphnia to rapid environmental change in a polluted lake (Jennifer Fox), and (10) the factors affecting food chain length in lakes of differing size and productivity (David Post; co-advised by Mike Pace).
Undergraduate students have worked in my lab either on my research projects (or those of my grad students and postdocs), or carry out their own independent studies. Independent investigations have involved (1) genes involved in DNA repair after UV damage in high mountain lake Daphnia (Paige Kulling) (2) predatory-prey dynamics in lab microcosms and theory by tuning the strength of rotifer predation on algae using salinity (Alex Looi, Kathryn Blackley, Dave Rosenberg), (3) strength of evolutionary rescue as a function of extent of inbreeding depression in rotifers (Kathryn Blackley), (4) rates of plankton dispersal among rock pools during rain-induced overflow events (Kara Pellowe), (5) tracing clonal lineages of algae using microsatellite-DNA to follow evolutionary response of algae to consumption by rotifers (Justin Meyer), (6) the patterns of Daphnia abundance in Austrian Alpine lakes with very long histories of fish introduction (Linda Perry Demma), (7) the identification of past invasions by exotic species in using molecular analysis of dormant eggs from lake sediments (Torrie Hanley), (8) molecular genetic analysis of genetic variation in copepod species with and without prolonged diapause, and genetic changes in algal prey during predator-prey cycles (Matt Holland). When appropriate, undergraduate students participate as co-authors on publications (e.g., Hewson, NG, Li, Aguirre, Looi et al. 2013; Meyer et al. 2006; Bohonak, Holland et al. 2006; Hairston...Perry et al. 2005; Duffy, Perry et al. 2000; Hairston, Perry et al. 1999 - all names except mine, Hewson’s, and Bohonak's were undergrads in my lab when the published work was carried out).
PAST RESEARCH
Eco-evolutionary dynamics and internally-driven temporal variation – Along with Stephen Ellner, Laura Jones (Cornell University), and a series of great postdocs (Gregor Fussmann, now on the faculty at McGill University; Takehito Yoshida, now on the faculty at Tokyo University; Lutz Becks, on the faculty at the University of Konstanz; Teppo Hiltunen, now on the faculty at the University of Turku; Brooks Miner, now on the faculty at Ithaca College, and Masato Yamamichi, now on the faculty at the University of Queensland), as well as a cadre of dedicated undergraduate students, we used highly controlled culture conditions to create predator-prey oscillations between algae and rotifers. We were able to predict the dynamics in our culture system with mechanistic mathematical models (Fussmann et al. 2000) and then to use these models to predict unexpected behaviors. One very interesting result was the observation that when the prey (algae) can evolve in response to temporally varying selection imposed by predation (rotifer grazing) and competition for limiting nutrients (nitrogen), the pattern of the predator-prey oscillations is radically altered (Yoshida et al. 2003). Under some conditions, "cryptic oscillations" of prey genotypes occur while total prey density remains essentially constant (Yoshida et al. 2007). Thus rapid contemporary evolution can dramatically alter what has traditionally been viewed as a purely ecological process (Becks et al. 2010). We have recently used this system to study the functional genomics of oscillating evolution of prey defense (cell clumping by Chlamydomonas) in response to predator-prey cycling (Becks et al. 2012).
In 2019, we published a paper (Schaffner & Govaert et al. 2019) showing that Daphnia mendotae evolves resistance to summer cyanobacterial blooms within a single season and that this evolution feeds back to affect the population dynamics of the Daphnia.

Microevolutionary processes – Predator-prey cycling is one of many temporally and spatially varying selection pressures. In evolutionary biology, I am interested in ways that organisms deal with variable environments and the way that rapid adaptive evolutionary response to environmental change (whether natural or anthropogenic) influences ecological dynamics. My colleagues and I (especially Monica Geber who dreamed up the approach, and Steve Ellner who has been instrumental in developing and applying it to our data) have devised methods for assessing the relative rates of evolutionary and ecological change (Hairston et al. 2005; Ellner et al. 2011). We estimate the magnitudes of the effect of both an evolving character and a changing environment on the rates of change of ecological processes.
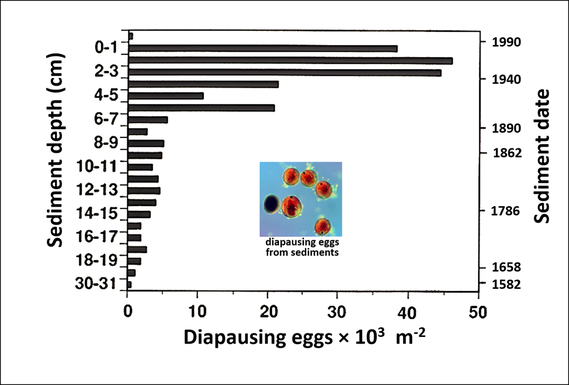
Dispersal between habitat patches can be of central importance for evolution in a spatially varying environment. Similarly, dispersal through time can be critical for evolution in an environment where selection fluctuates temporally (Ellner and Hairston, 1994; Hairston et al. 1996). This temporal dispersal is created in the crustacean zooplankton that I study through long-lived dormant eggs (see figure above). These eggs routinely survive for decades in lake sediments (Hairston 1996), and in extreme instances, for centuries (Hairston et al. 1995). In the case of the copepod Onychodiaptomus sanguineus living in small lakes in Rhode Island, an important fitness character is the seasonal timing of the production of dormant eggs. In summer, the animals would normally be subjected to intense predation by fish, but the copepods avoid this by producing dormant eggs that rest on the lake bottom where the fish do not find them. The optimal time in spring to begin dormant-egg production varies from year to year depending upon fish density and activity. There is genetic variation for the timing of dormancy (Hairston and Dillon 1990), and the populations respond to changes between years in selection pressure (Hairston and De Stasio 1988; Hairston and Dillon 1990). Theory suggests that this fluctuating selection, combined with the overlap in generations created by the dormant "egg bank," creates conditions for the maintenance of genetic variation for the seasonal timing of dormancy. It is this genetic variation that permits response to changes in natural selection. Interestingly, the variation maintained is not continuous. Rather, two distinct phenotypes coexist (Hairston et al. 1996; Ellner et al. 1998). One deals with year-to-year variations in fish predation by switching to dormancy early in the season (thus avoiding even the earliest onset of predation). Copepods with an early switch also have a high probability of hatching from the egg bank. The other phenotype suffers low fitness in most years, but in years of late onset of predation can have very high fitness. These late-switching copepods have a relatively low probability of hatching from dormancy in the sediments. The exact diapause timing by the copepod population in any given year is the result of past natural selection combined with the influence of proportions of two phenotypes hatching from the lake sediments (Ellner et al. 1999). Based on the method in Hairston et al. (2005), the rate of adaptive evolution in this system is on the order of 20% of the rate of ecological change.
Dispersal between habitat patches can be of central importance for evolution in a spatially varying environment. Similarly, dispersal through time can be critical for evolution in an environment where selection fluctuates temporally (Ellner and Hairston, 1994; Hairston et al. 1996). This temporal dispersal is created in the crustacean zooplankton that I study through long-lived dormant eggs (see figure above). These eggs routinely survive for decades in lake sediments (Hairston 1996), and in extreme instances, for centuries (Hairston et al. 1995). In the case of the copepod Onychodiaptomus sanguineus living in small lakes in Rhode Island, an important fitness character is the seasonal timing of the production of dormant eggs. In summer, the animals would normally be subjected to intense predation by fish, but the copepods avoid this by producing dormant eggs that rest on the lake bottom where the fish do not find them. The optimal time in spring to begin dormant-egg production varies from year to year depending upon fish density and activity. There is genetic variation for the timing of dormancy (Hairston and Dillon 1990), and the populations respond to changes between years in selection pressure (Hairston and De Stasio 1988; Hairston and Dillon 1990). Theory suggests that this fluctuating selection, combined with the overlap in generations created by the dormant "egg bank," creates conditions for the maintenance of genetic variation for the seasonal timing of dormancy. It is this genetic variation that permits response to changes in natural selection. Interestingly, the variation maintained is not continuous. Rather, two distinct phenotypes coexist (Hairston et al. 1996; Ellner et al. 1998). One deals with year-to-year variations in fish predation by switching to dormancy early in the season (thus avoiding even the earliest onset of predation). Copepods with an early switch also have a high probability of hatching from the egg bank. The other phenotype suffers low fitness in most years, but in years of late onset of predation can have very high fitness. These late-switching copepods have a relatively low probability of hatching from dormancy in the sediments. The exact diapause timing by the copepod population in any given year is the result of past natural selection combined with the influence of proportions of two phenotypes hatching from the lake sediments (Ellner et al. 1999). Based on the method in Hairston et al. (2005), the rate of adaptive evolution in this system is on the order of 20% of the rate of ecological change.
{kind=link}
In a separate study of the Daphnia galeata in Lake Constance (Europe), colleagues at the Max-Planck-Institute for Limnology (now the Max-Planck-Institute for Evolutionary Biology), Plön, Germany, and from my lab at Cornell, used individuals hatched from diapausing eggs extracted from the lake sediments of different time periods to show how the population evolved resistance to dietary cyanobacteria during the course of lake eutrophication (Hairston et al. 1999, 2001). Using reaction norms, we have gone on to show that the degree of phenotypic plasticity evolved in this population as cyanobacteria became more prevalent in the plankton during the period 1960 - 1980. The Daphnia genotypes taken from sediments deposited during the 1960s show a significantly larger average change in performance (specific juvenile growth rate) than do genotypes obtained from sediments dated from the late 1970s (Hairston and De Meester 2008).
Most recently, we have shown that Daphnia mendotae in Oneida Lake, site of the Cornell Biological Field Station, evolves resistance to cyanobacteria over the course of a single season (about 15 Daphnia generations), and that this sets up an eco-evolutionary dynamic in which grazer evolution can influence seasonal plankton dynamics (L. Schaffner & Govaert et al. 2019).
Most recently, we have shown that Daphnia mendotae in Oneida Lake, site of the Cornell Biological Field Station, evolves resistance to cyanobacteria over the course of a single season (about 15 Daphnia generations), and that this sets up an eco-evolutionary dynamic in which grazer evolution can influence seasonal plankton dynamics (L. Schaffner & Govaert et al. 2019).
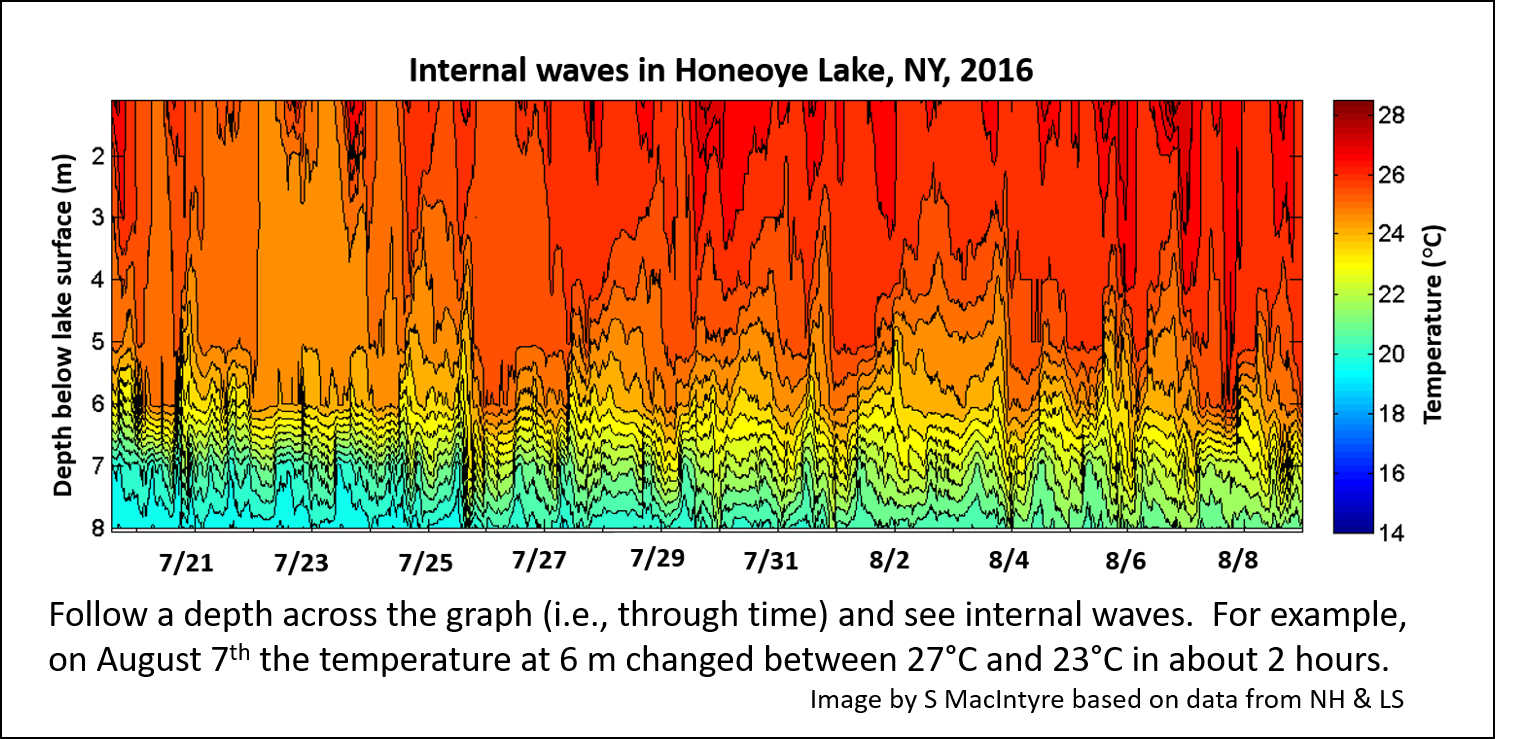
Causes of cyanobacterial blooms in an unpolluted lake – Current lab research continues after my retirement on the causes of blooms of cyanobacteria in a beautiful small Finger Lake – Honeoye Lake. Every summer there are dense occurrences of cyanobacterial scums even though the lake watershed is nearly completely forested and all houses along the shore have been connected to a sewer system that carries waste away from the lake. Our working hypothesis is that nutrients buried in the sediments of the lake are a legacy from past pollution. These nutrients are released into the hypolimnion in summer, but the shallow depth of the lake makes mixing of the nutrients into the surface waters, where they stimulate phytoplankton growth, fairly common. Our data suggest (and detailed modeling is underway to test the idea) that even without complete turnover of the water column, internal waves and upwelling events along the thermocline are sufficient to enrich the surface waters. Wouldn’t it be interesting if physical mixing processes in lakes ultimately drives the evolution of Daphnia via the stimulation of cyanobacterial blooms?


Community dynamics – At the community level, long-term dormancy may provide a mechanism for maintaining species diversity in a changing environment. At Onondaga Lake, New York State, industrial development created a lake environment that changed dramatically over the past century. I and others in my lab have studied the ability of the zooplankton community to respond to these changes through reintroduction from the egg bank, and the possible effects of pollution in the sediments on reducing the viability of dormant eggs (and hence reducing community response). One interesting result is the replacement of native species of Daphnia by exotic species during the period of peak pollution (by salt waste, heavy metal inputs, and nutrient pollution). The exotic species (D. exilis and D. curvirostris) are themselves native to saline environments in the southwestern U.S. and Europe (Hairston et al. 1999, Duffy et al. 2000).
A group of people from my lab have used molecular markers to explore the genetic changes in populations over time through the analysis of dormant eggs. One application has been studies rates of microevolutionary change. We used allozymes on animals hatched from the sediments to show that Onondaga Lake was invaded by D. exilis through the hatching of a single egg (genotype). Analysis of the 12S region of mtDNA extracted directly from diapausing eggs was used to identify the invasion of Onondaga Lake during the 1950s by D. curvirostris when the eggs could not be hatched (Duffy et al. 2000). Another use of molecular techniques is in understanding the frequency of reestablishment of populations from the egg bank, in comparison to reestablishment via dispersal from outside the lake. Together with colleagues at the Max-Planck-Institute, Andy Bohonak and Matt Holland from my lab used the COI region of mtDNA in copepods to evaluate the role of prolonged diapause in the nature of genetic variation in two species of diaptomid copepods (Bohonak et al. 2006). Recently Ian Hewson in collaboration with my lab identified viruses of Daphnia: we found that host death rates correlate, seasonally with viral prevalence and abundance.
A group of people from my lab have used molecular markers to explore the genetic changes in populations over time through the analysis of dormant eggs. One application has been studies rates of microevolutionary change. We used allozymes on animals hatched from the sediments to show that Onondaga Lake was invaded by D. exilis through the hatching of a single egg (genotype). Analysis of the 12S region of mtDNA extracted directly from diapausing eggs was used to identify the invasion of Onondaga Lake during the 1950s by D. curvirostris when the eggs could not be hatched (Duffy et al. 2000). Another use of molecular techniques is in understanding the frequency of reestablishment of populations from the egg bank, in comparison to reestablishment via dispersal from outside the lake. Together with colleagues at the Max-Planck-Institute, Andy Bohonak and Matt Holland from my lab used the COI region of mtDNA in copepods to evaluate the role of prolonged diapause in the nature of genetic variation in two species of diaptomid copepods (Bohonak et al. 2006). Recently Ian Hewson in collaboration with my lab identified viruses of Daphnia: we found that host death rates correlate, seasonally with viral prevalence and abundance.

Visual ability and diet choice by fish – Another interest of mine has been the way that zooplanktivorous fish locate their prey and how they make dietary decisions. William Walton (now at UC Riverside), Stephen Easter (U Michigan), and I studied how the ability of bluegill sunfish (Lepomis machrochirus) to detect prey visually increases during ontogeny and the effect that this has on diet choice (Hairston et al. 1982, Walton et al. 1992). As fish grow, sensory cells are added to the retina causing visual resolution to increase. At the same time, the eye grows larger and the retina recedes from the lens, with the result that visual resolution declines. The net effect is a non-linear increase in visual resolution as fish grow. Behavioral studies show that the feeding reaction (orientation and capture) of fish to their zooplankton prey follow this non-linear pattern closely. The ability of fish to find their prey increases with age (and size), and this in concert with changing ability to handle prey after they are captured results is a diet choice that closely matches predictions of optimal foraging theory.

Interactions between community composition and nutrient cycling – With David Hambright (now at U. Oklahoma), Robert Howarth and William Schaffner (Cornell EEB), we have studied the role that zooplankton community structure has on the ecosystem-level processes of nutrient cycling. Using the Cornell Experimental Ponds Facility, we manipulated zooplankton community structure by having fish present (produces a copepod-dominated community) or absent (produces a Daphnia-dominated community). We also manipulated nutrient additions with all ponds having high phosphorous inputs, but some having low nitrogen additions and others having high nitrogen additions. Based on research by others, we would have expected high rates of nitrogen fixation in ponds with low N:P ratios. However, we found this result only in the presence of fish (where grazing by zooplankton is relatively low). When fish are absent, the Daphnia graze down the cyanobacteria to low levels and N-fixation is greatly reduced (Hambright et al. 2007a,b). Although cyanobacteria are generally considered poor food for Daphnia, these grazers appear early in the year and prevent any cyanobacterial bloom from starting (Schaffner et al. 1994).